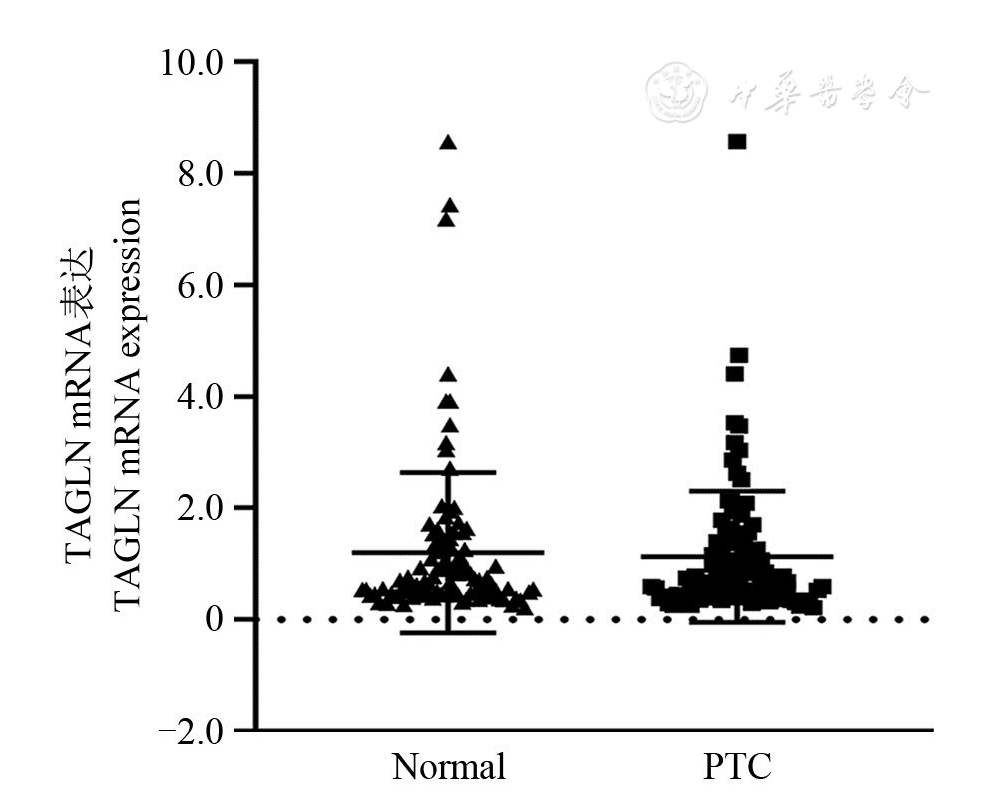
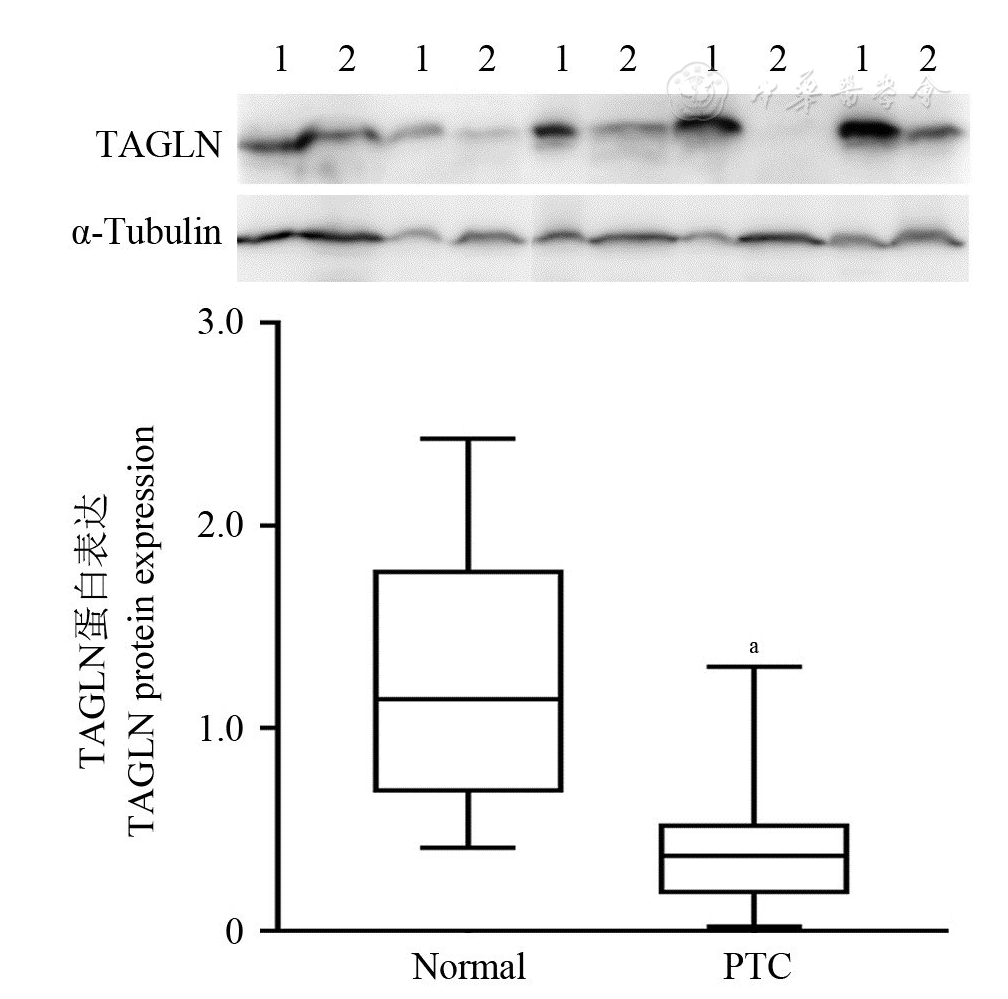
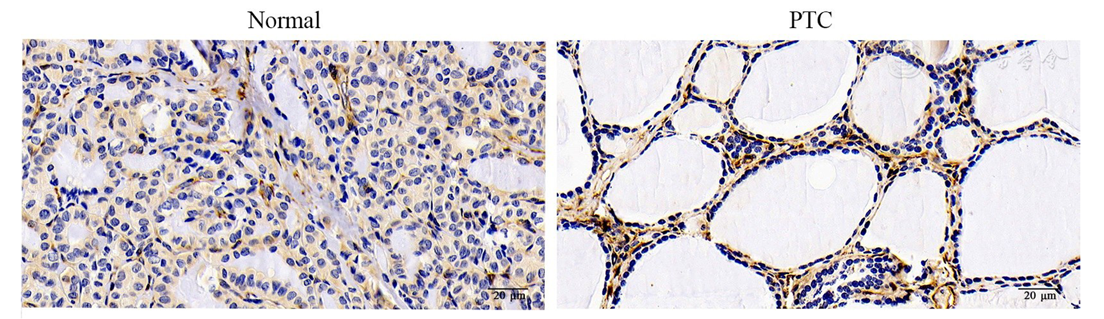
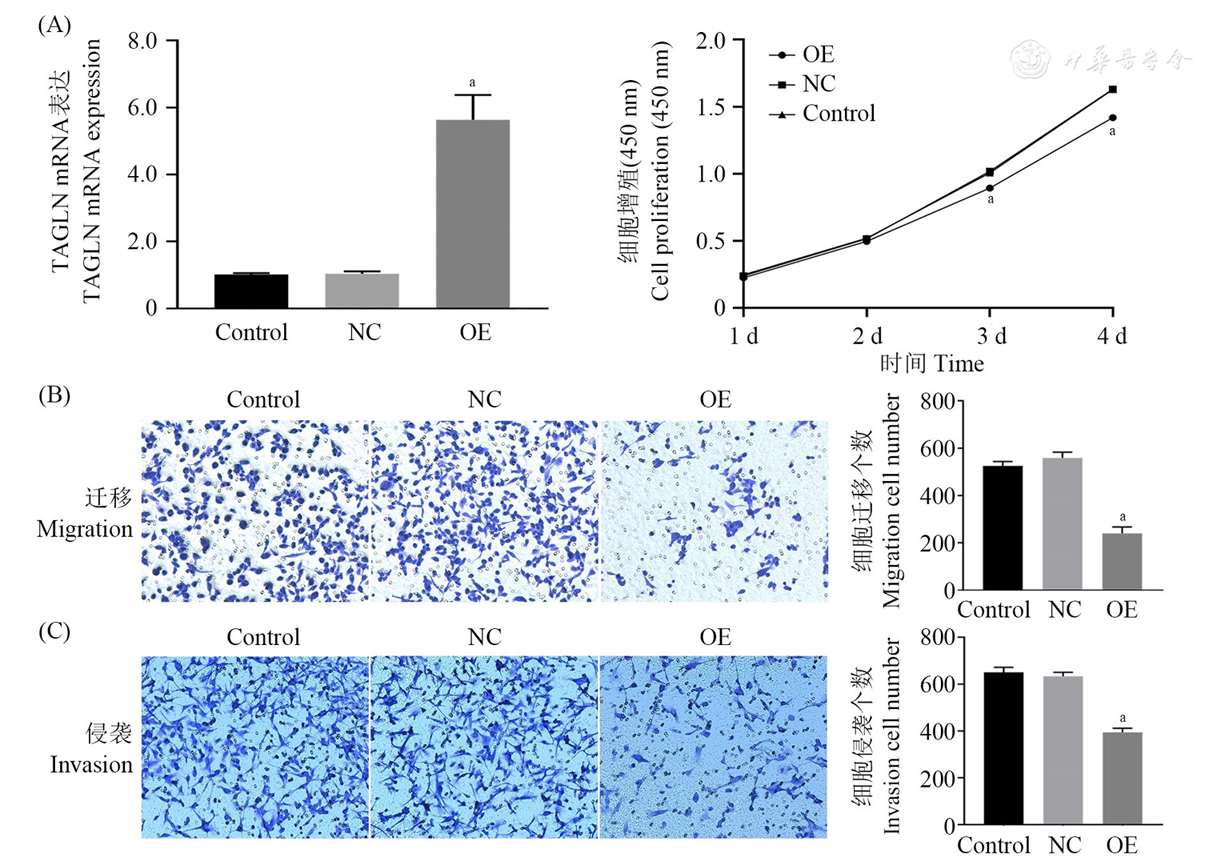
参考文献
[1] Xing M. Molecular pathogenesis and mechanisms of thyroid cancer[J]. Nat Rev Cancer, 2013, 13(3): 184-199. DOI: 10.1038/nrc3431.
[2] Roman BR, Morris LG, Davies L. The thyroid cancer epidemic, 2017 perspective[J]. CurrOpin Endocrinol Diabetes Obes, 2017, 24(5): 332-336. DOI: 10.1097/MED.0000000000000359.
[3] Cabanillas ME, McFadden DG, Durante C. Thyroid cancer[J]. Lancet, 2016, 388(10061): 2783-2795. DOI: 10.1016/S0140-6736(16)30172-6.
[4] Ambrosi F, Righi A, Ricci C, et al. Hobnail variant of papillary thyroid carcinoma: a literature review[J]. Endocr Pathol, 2017, 28(4): 293-301. DOI: 10.1007/s12022-017-9502-7.
[5] Chakraborty R, Saddouk FZ, Carrao AC, et al. Promoters to study vascular smooth muscle[J]. Arterioscler Thromb Vasc Biol, 2019, 39(4): 603-612. DOI: 10.1161/ATVBAHA.119.312449.
[6] Dvorakova M, Nenutil R, Bouchal P. Transgelins, cytoskeletal proteins implicated in different aspects of cancer development[J]. Expert Rev Proteomics, 2014, 11(2): 149-165. DOI: 10.1586/14789450.2014.860358.
[7] Pawlak G, Helfman DM. Cytoskeletal changes in cell transformation and tumorigenesis[J]. CurrOpin Genet Dev, 2001, 11(1): 41-47. DOI: 10.1016/s0959-437x(00)00154-4.
[8] Dong LH, Wen JK, Liu G, et al. Blockade of the Ras-extracellular signal-regulated kinase 1/2 pathway is involved in smooth muscle 22 alpha-mediated suppression of vascular smooth muscle cell proliferation and neointima hyperplasia[J]. Arterioscler Thromb Vasc Biol, 2010, 30(4): 683-691. DOI: 10.1161/ATVBAHA.109.200501.
[9] Siegel RL, Miller KD, Jemal A. Cancer statistics, 2019[J]. CA Cancer J Clin, 2019, 69(1): 7-34. DOI: 10.3322/caac.21551.
[10] Morris LG, Tuttle RM, Davies L. Changing trends in the incidence of thyroid cancer in the United States[J]. JAMA Otolaryngol Head Neck Surg, 2016, 142(7): 709-711. DOI: 10.1001/jamaoto.2016.0230.
[11] Borrelli N, Panebianco F, Condello V, et al. Characterization of activ-ating mutations of the MEK1 gene in papillary thyroid carcinomas[J]. Thyroid, 2019, 29(9): 1279-1285. DOI: 10.1089/thy.2019.0065.
[12] Li M, Li S, Lou Z, et al. Crystal structure of human transgelin[J]. J Struct Biol, 2008, 162(2): 229-236. DOI: 10.1016/j.jsb.2008.01.005.
[13] Hirata H, Gupta M, Vedula SR, et al. Quantifying tensile force and ERK phosphorylation on actin stress fibers[J]. Methods Mol Biol, 2017, 1487: 223-234. DOI: 10.1007/978-1-4939-6424-6_16.
[14] Tsui KH, Lin YH, Chang KS, et al. Transgelin, a p53 and PTEN-upregulated gene, inhibits the cell proliferation and invasion of human bladder carcinoma cells in vitro and in vivo[J]. Int J Mol Sci, 2019, 20(19): 4946. DOI: 10.3390/ijms20194946.
[15] Yeo M, Park HJ, Kim DK, et al. Loss of SM22 is a characteristic signature of colon carcinogenesis and its restoration suppresses colon tumorigenicity in vivo and in vitro[J]. Cancer, 2010, 116(11): 2581-2589. DOI: 10.1002/cncr.25003.
[16] Zhang J, Wang K, Zhang J, et al. Using proteomic approach to identify tumor-associated proteins as biomarkers in human esophageal squamous cell carcinoma[J]. J Proteome Res, 2011, 10(6): 2863-2872. DOI: 10.1021/pr200141c.
[17] Wu X, Dong L, Zhang R, et al. Transgelin overexpression in lung adenocarcinoma is associated with tumor progression[J]. Int J Mol Med, 2014, 34(2): 585-591. DOI: 10.3892/ijmm.2014.1805.
[18] Mahal BA, Alshalalfa M, Zhao SG, et al. Genomic and clinical charac-terization of stromal infiltration markers in prostate cancer[J]. Cancer, 2020, 126(7): 1407-1412. DOI: 10.1002/cncr.32688.
[19] Zadeh Fakhar HB, Zali H, Rezaie-Tavirani M, et al. Proteome profiling of low grade serous ovarian cancer[J]. J Ovarian Res, 2019, 12(1): 64. DOI: 10.1186/s13048-019-0535-z.
[20] Petrulea MS, Plantinga TS, Smit JW, et al. PI3K/Akt/mTOR: A promising therapeutic target for non-medullary thyroid carcinoma[J]. Cancer Treat Rev, 2015, 41(8): 707-713. DOI: 10.1016/j.ctrv.2015.06.005.
[21] Milosevic Z, Pesic M, Stankovic T, et al. Targeting RAS-MAPK-ERK and PI3K-AKT-mTOR signal transduction pathways to chemosensitize anaplastic thyroid carcinoma[J]. Transl Res, 2014, 164(5): 411-423. DOI: 10.1016/j.trsl.2014.06.005.
[22] Zou X, Blank M. Targeting p38 MAP kinase signaling in cancer through post-translational modifications[J]. Cancer Lett, 2017, 384: 19-26. DOI: 10.1016/j.canlet.2016.10.008.