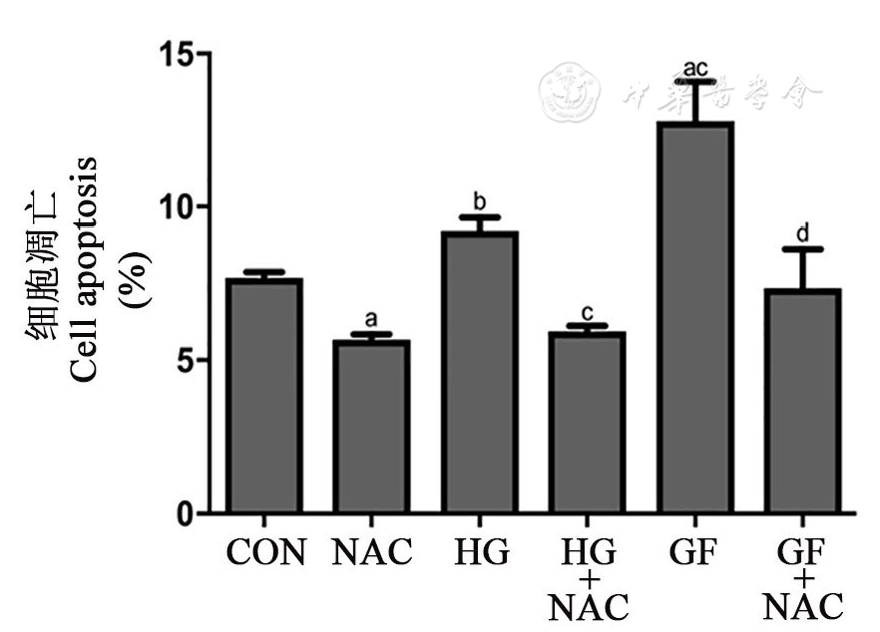
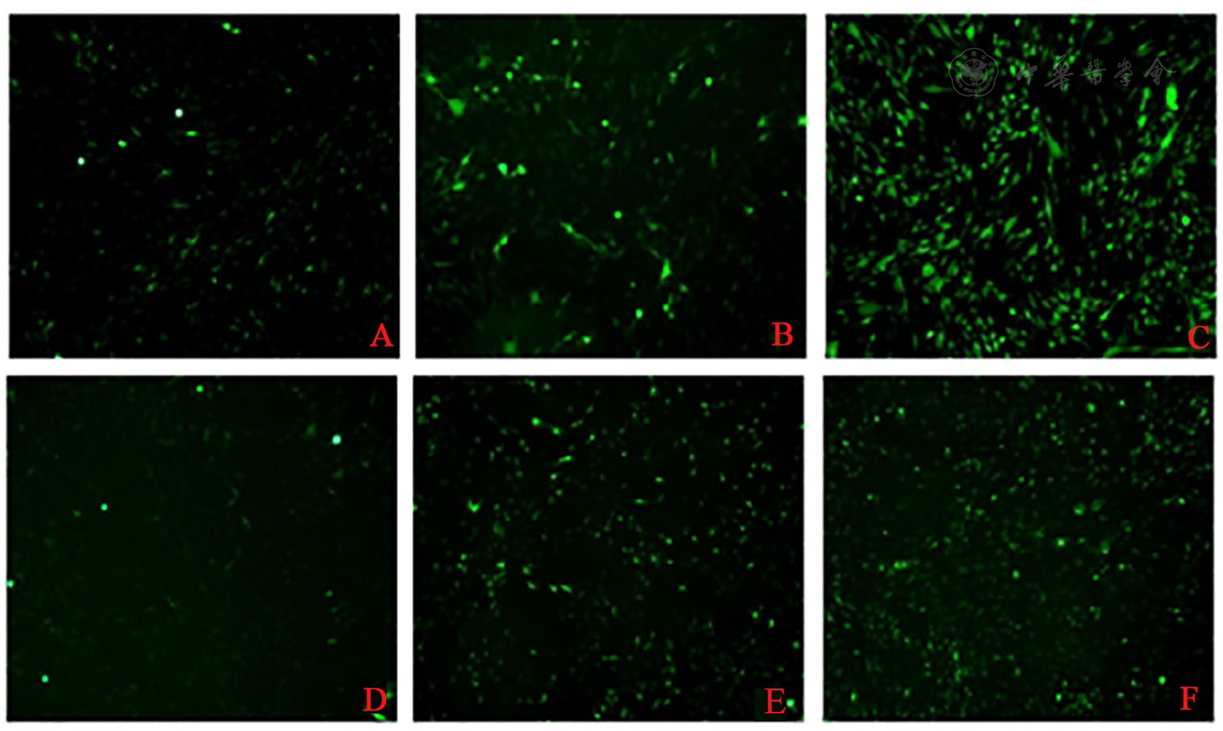
文献
[1] Oud L, Spellman C. The association of glucose variability and home discharge among survivors of critical illness managed with a computerized decision-support tool for glycemic control [J]. J Diabetes Sci Technol, 2014,8(2):277-285. DOI: 10.1177/1932296813518136.
[2] Giacco F, Brownlee M. Oxidative stress and diabetic complications[J]. Circ Res, 2010,107(9):1058-1070. DOI: 10.1161/CIRCRESAHA.110.223545.
[3] Yamagishi S. Role of advanced glycation end products(AGEs)in osteoporosis in diabetes[J]. Curr Drug Targets, 2011,12(14):2096-2102. DOI: 10.2174/138945011798829456.
[4] Kim MK, Jung HS, Yoon CS, et al. The effect of glucose fluctuation on apoptosis and fluctuation of INS-1 pancreatic beta cells[J]. Korean Diabetes J, 2010,34(1):47-54. DOI: 10.4093/kdj.2010.34.1.47.
[5] Wu N, Shen H, Liu H, et al. Acute blood glucose fluctuation enhances rat aorta endothelial cell apoptosis, oxidative stress and pro-inflammatory cytokine expression in vivo[J]. Cardiovasc Diabetol, 2016,15(1):109. DOI: 10.1186/s12933-016-0427-0.
[6] Sun LQ, Xue B, Lin XJ, et al. Inhibitory effects of salvianolic acid B on apoptosis of Schwann cells and its mechanism induced by intermittent high glucose[J]. Life Sci, 2012,90(3-4):99-108. DOI: 10.1016/j.lfs.2011.10.001.
[7] 王小菊,冯正平,李济伶,等. 氧化应激在高糖诱导成骨细胞凋亡中的作用[J]. 重庆医科大学学报,2015,40(7):960-964. DOI: 10.13406/j.cnki.cyxb.000696.
[8] Monnier L, Mas E, Ginet C, et al. Activation of oxidative stress by acute glucose fluctuations compared with sustained chronic hyperglycemia in patients with type 2 diabetes[J]. JAMA, 2006,295(14):1681-1687. DOI: 10.1001/jama.295.14.1681.
[9] Meng X, Gong C, Cao B, et al. Glucose fluctuations in association with oxidative stress among children with T1DM: comparison of different phases[J]. J Clin Endocrinol Metab, 2015,100(5):1828-1836. DOI: 10.1210/jc.2014-2879.
[10] Xia J, Qu Y, Yin C, et al. Optical coherence tomography assessment of glucose fluctuation impact on the neointimal proliferation after stent implantation in a diabetic/hypercholesterolemic swine model[J]. Int Heart J, 2017,58(4):608-614. DOI: 10.1536/ihj.16-574.
[11] Wu N, Shen H, Wang Y, et al. Role of the PKCβII/JNK signaling pathway in acute glucose fluctuation-induced apoptosis of rat vascular endothelial cells[J]. Acta diabetologica, 2017,54(8):727-736. DOI: 10.1007/s00592-017-0999-5
[12] Ducy P, Schinke T, Karsenty G. The osteoblast: a sophisticated fibroblast under central surveillance[J]. Science, 2000,289(5484):1501-1504. DOI: 10.1126/science.289.5484.1501.
[13] Massé PG, Pacifique MB, Tranchant CC, et al. Bone metabolic abnormalities associated with well-controlled type 1 diabetes(IDDM)in young adult women: a disease complication often ignored or neglected[J]. J Am Coll Nutr, 2010,29(4):419-429. DOI: 10.1080/07315724.2010.10719859.
[14] Bai XC, Lu D, Bai J, et al. Oxidative stress inhibits osteoblastic differentiation of bone cells by ERK and NF-kappaB[J]. Biochem Biophys Res Commun, 2004,314(1):197-207. DOI: 10.1016/j.bbrc.2003.12.073.
[15] Fatokun AA, Stone TW, Smith RA. Hydrogen peroxide-induced oxidative stress in MC3T3-E1 cells: The effects of glutamate and protection by purines[J]. Bone, 2006,39(3):542-551. DOI: 10.1016/j.bone.2006.02.062.
[16] Wang X, Feng Z, Li J, et al. High glucose induces autophagy of MC3T3-E1 cells via ROS-AKT-mTOR axis[J]. Mol Cell Endocrinol, 2016,429:62-72. DOI: 10.1016/j.mce.2016.03.036.
[17] 周玮,万利,张勇,等. 高糖及高糖波动对MG63细胞株氧化应激的影响[J]. 中国骨质疏松杂志,2011,12(4):1048-1051. DOI: 10.3969/j.issn.1006-7108.2011.12.004.
[18] 陆卫平,周玮. 不同浓度葡萄糖对MG63细胞增殖和细胞周期的影响[J]. 南京医科大学学报(自然科学版), 2010,30(9):1255-1257.
[19] Feng YL, Tang XL. Effect of glucocorticoid-induced oxidative stress on the expression of Cbfa1[J]. Chem Biol Interact, 2014,207:26-31. DOI: 10.1016/j.cbi.2013.11.004.
[20] Wauquier F, Leotoing L, Coxam V, et al. Oxidative stress in bone remodeling and disease[J]. Trends Mol Med, 2009,15(10):468-477. DOI: 10.1016/j.molmed.2009.08.004.
[21] Liang D, Xiang L, Yang M, et al. ZnT7 can protect MC3T3-E1 cells from oxidative stress-induced apoptosis via PI3K/Akt and MAPK/ ERK signaling pathways[J].Cell Signal, 2013,25(5):1126-1135. DOI: 10.1016/j.cellsig.2013.02.003.
[22] 王秉义,潘剑. 丹参素拮抗氧化应激所致骨质疏松并通过PI3k/Akt通路减少成骨细胞的凋亡[J]. 中国骨质疏松杂志,2017,23(1):1-5. DOI: 10.3969/j.issn.1006-7108.2017.01.001.
[23] 付卓栋,付超,孙慧君,等. 硫辛酸对体外培养的MC3T3-E1成骨细胞骨形成的影响[J]. 中华骨质疏松与骨矿盐疾病杂志,2016,9(1):43-51. DOI: 10.3969/j.issn.1674-2591.2016.01.008.
[24] 郭宝磊,杨茂伟,梁单,等. 高糖通过提高ROS水平和钙超载诱导小鼠MC3T3-E1成骨细胞凋亡[J]. 中国病理生理杂志,2012,28(2):292-297. DOI: 10.3969/j.issn.1000-4718.2012.02.019.