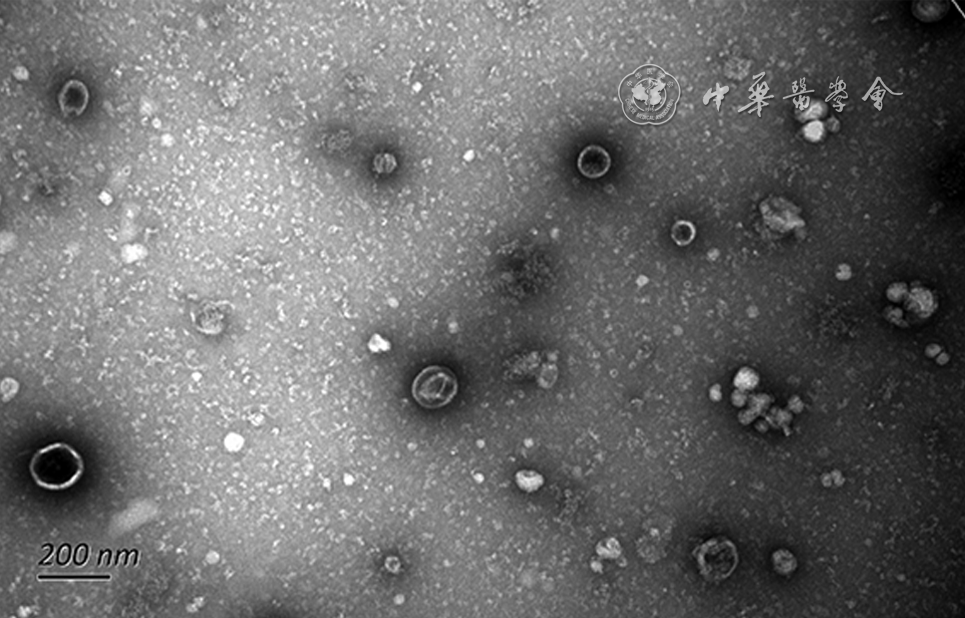
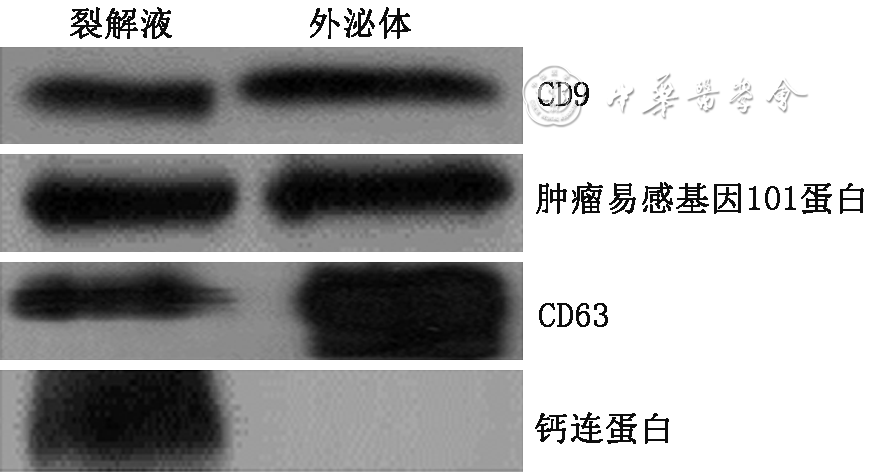
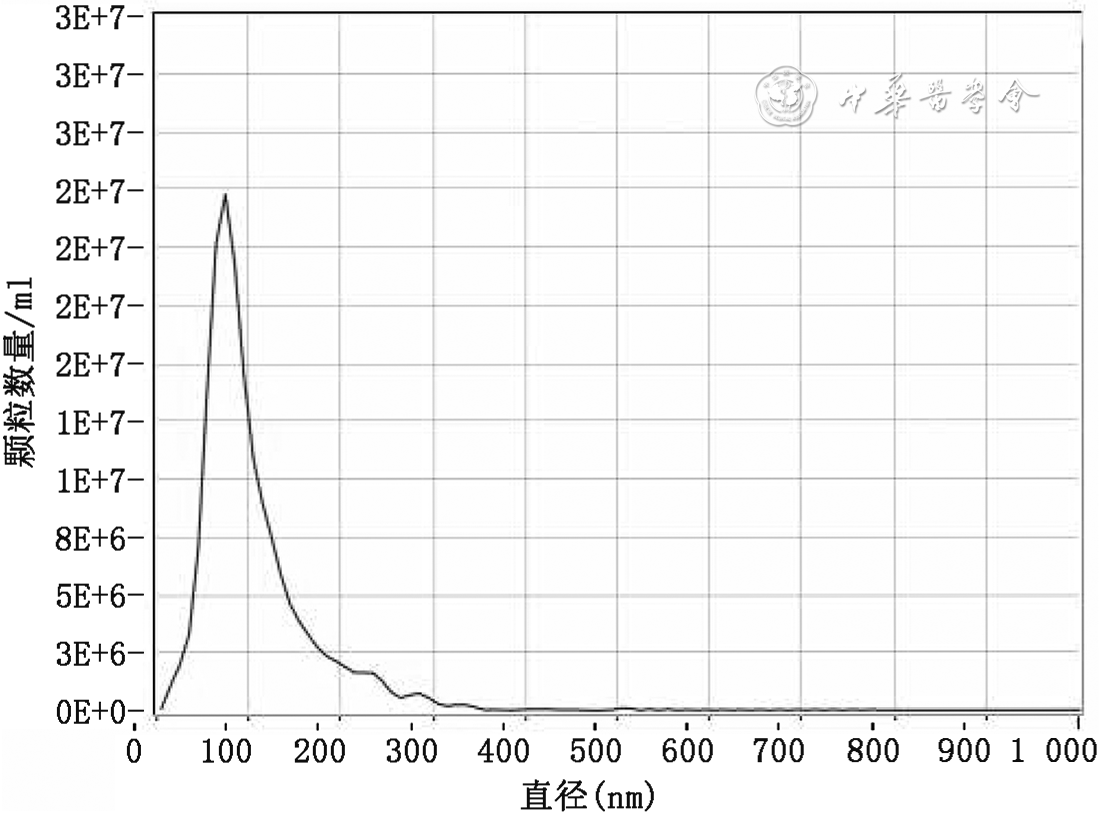
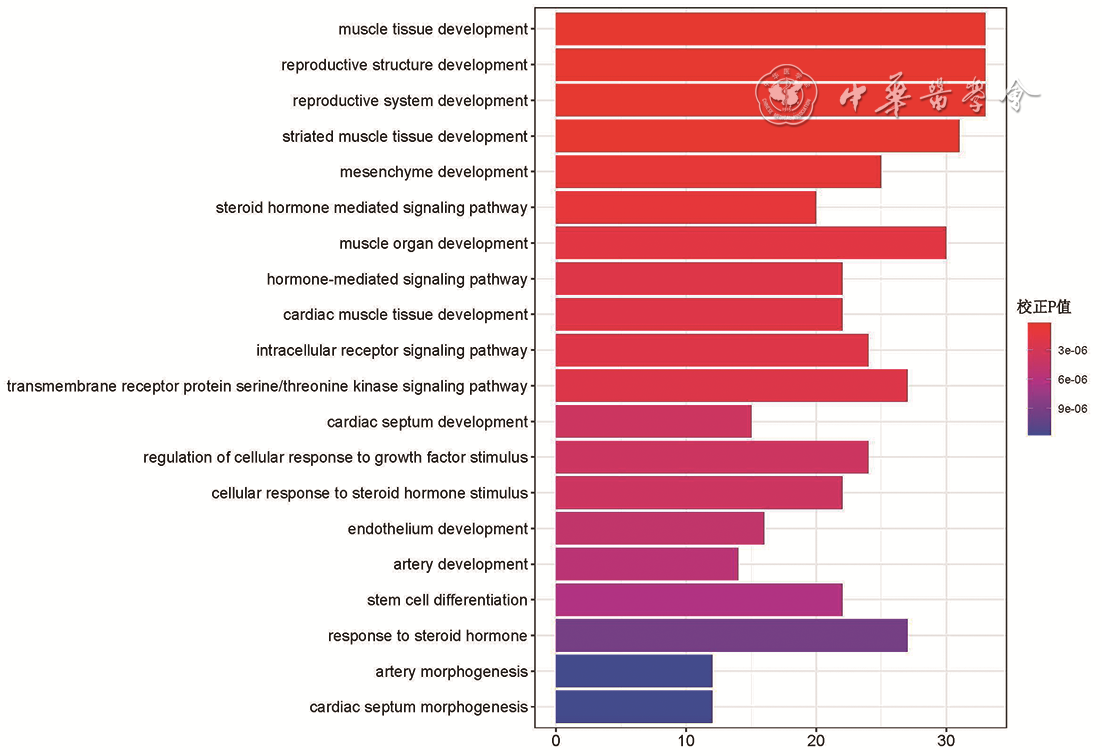
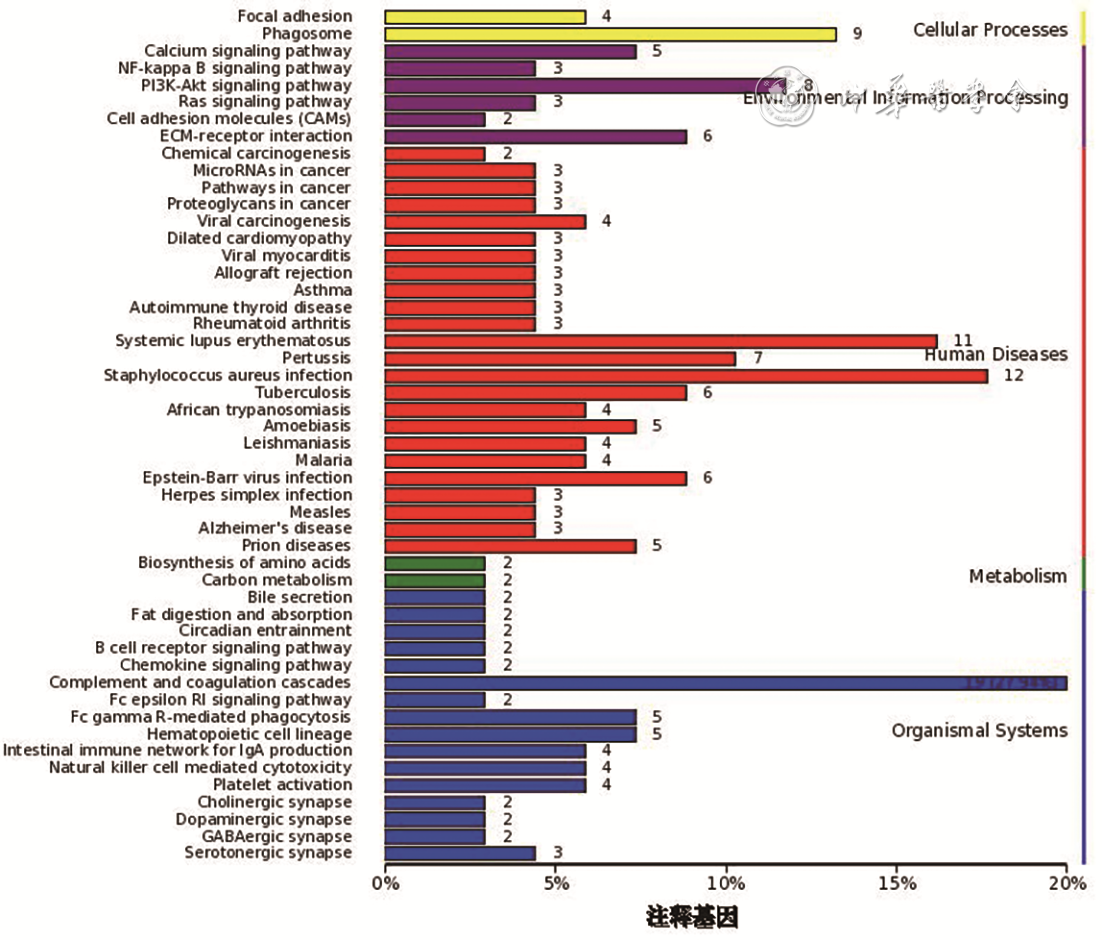
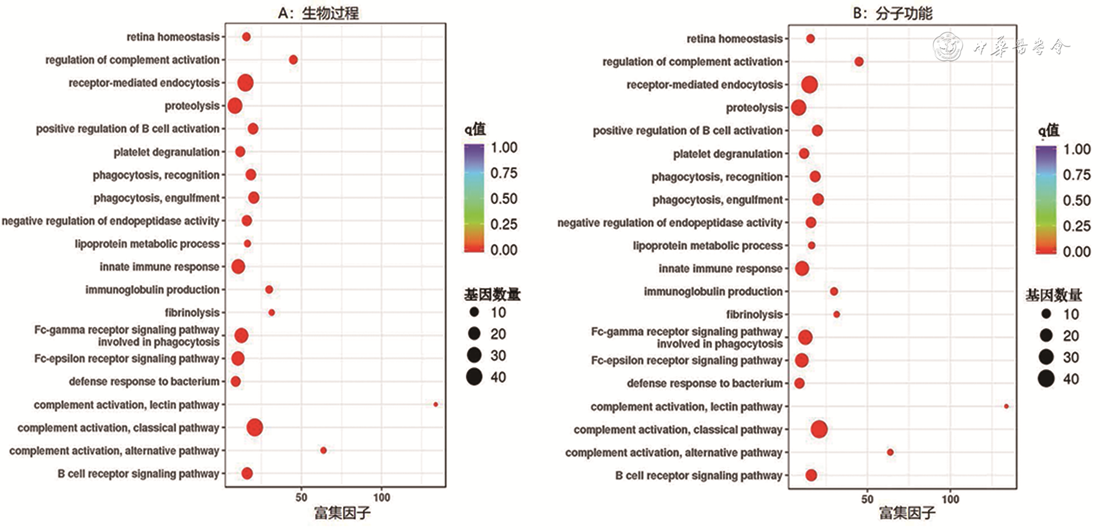
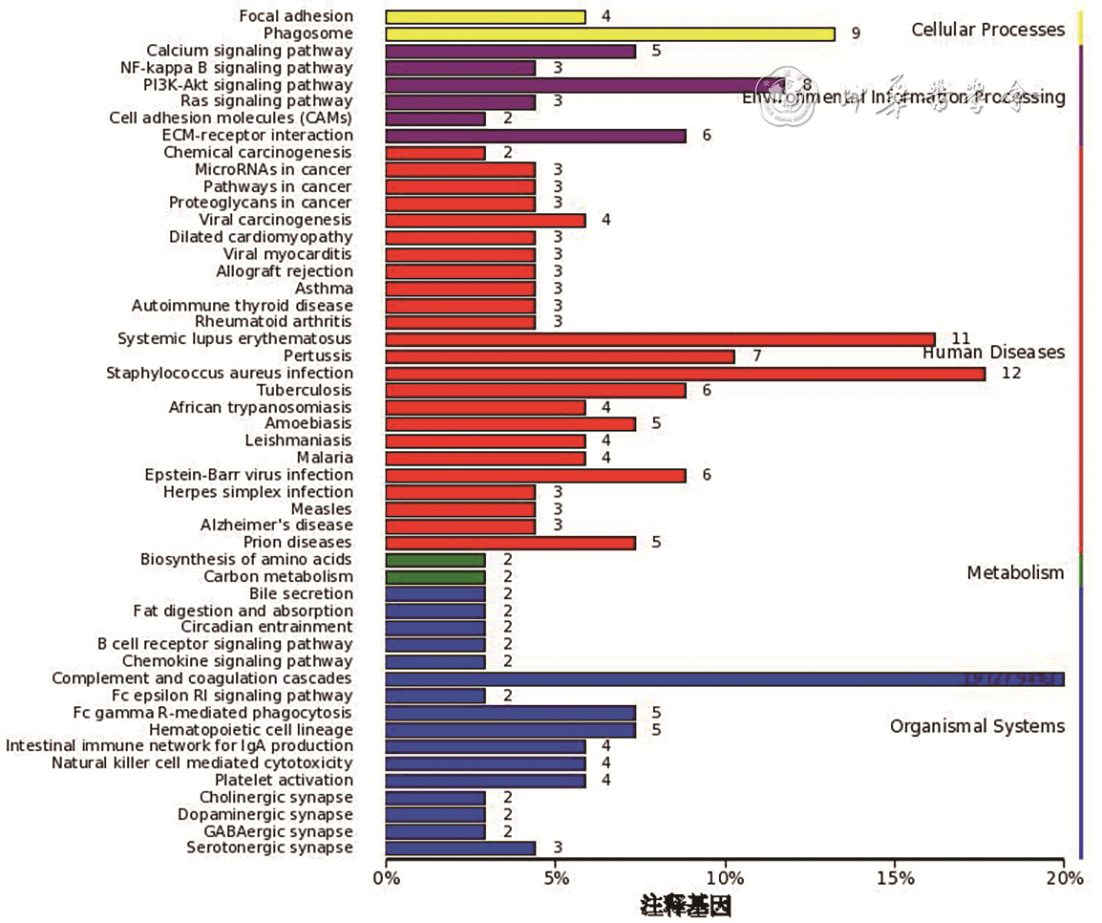
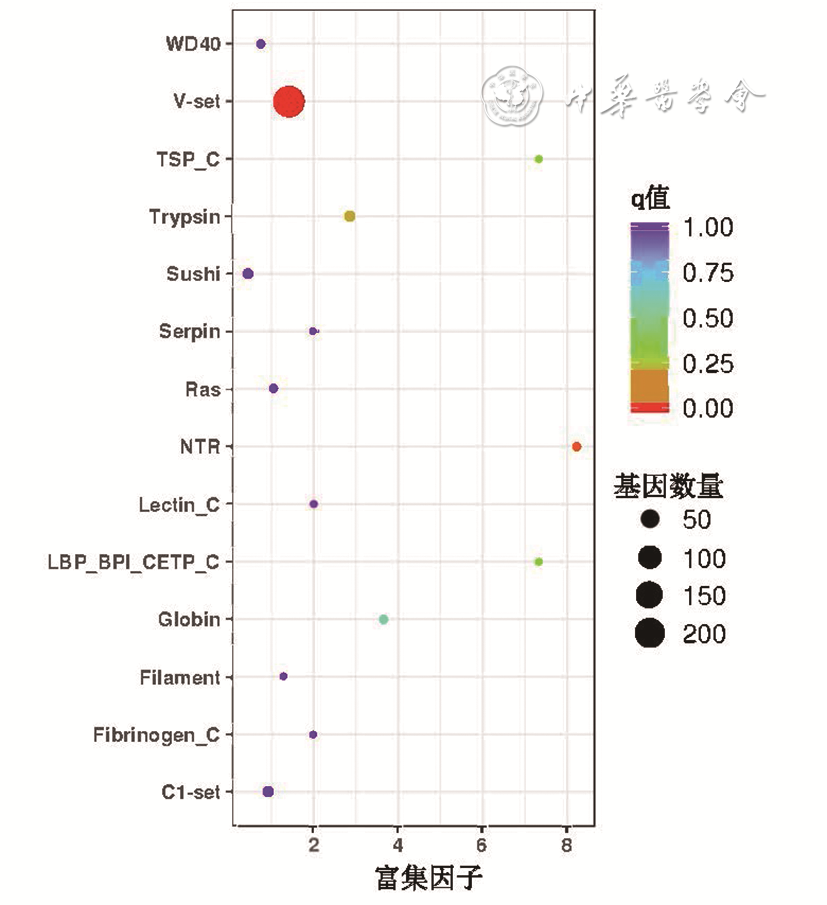
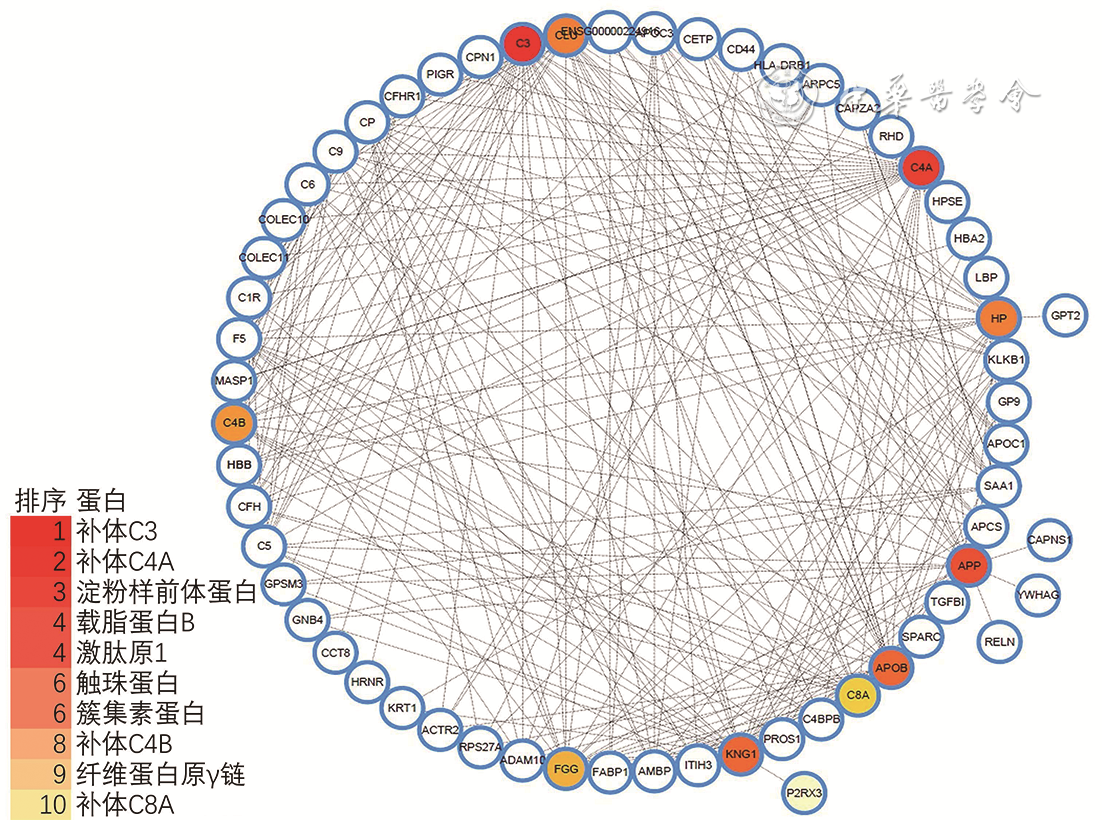
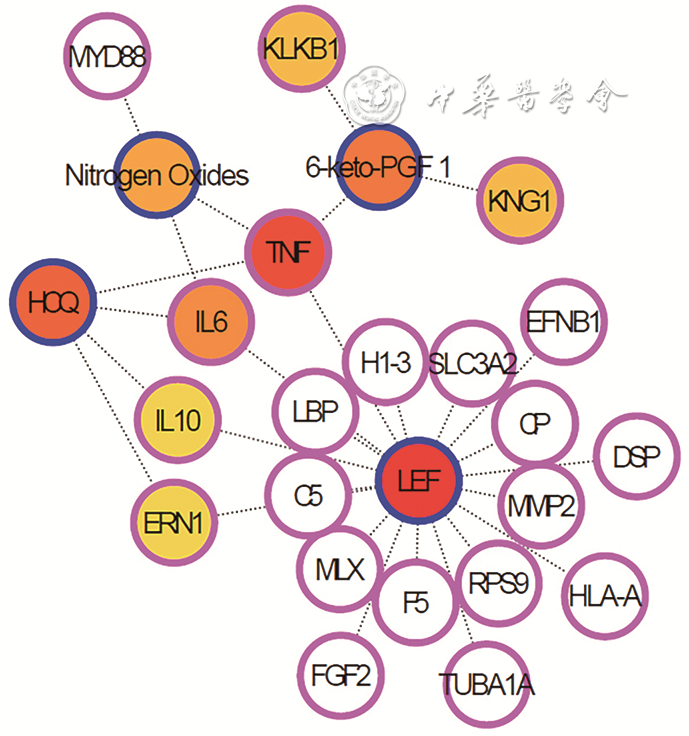
:黄碧青, 李菁, 田新平, 等. 多发性大动脉炎患者血浆外泌体携带的差异微小RNA和蛋白表达谱分析[J]. 中华内科杂志, 2023, 62(1): 61-69. DOI: 10.3760/cma.j.cn112138-20220208-00096.利益冲突:所有作者声明无利益冲突参考文献
[1] Zaldivar Villon MLF, de la Rocha JAL, Espinoza LR. Takayasu arteritis: recent developments[J]. Curr Rheumatol Rep, 2019, 21(9):45. DOI: 10.1007/s11926-019-0848-3.
[2] Wu X, Liu Y, Wei W, et al. Extracellular vesicles in autoimmune vasculitis-little dirts light the fire in blood vessels[J]. Autoimmun Rev, 2019, 18(6):593-606. DOI: 10.1016/j.autrev.2018.12.007.
[3] Shah D, Das P, Alam MA, et al. MicroRNA-34a promotes endothelial dysfunction and mitochondrial-mediated apoptosis in murine models of acute lung injury[J]. Am J Respir Cell Mol Biol, 2019, 60(4):465-477. DOI: 10.1165/rcmb.2018-0194OC.
[4] Zhang FM, Zheng WH, Wang HJ. MiR-34a-5p inhibition attenuates LPS-induced endothelial cell injury by targeting FOXM1[J]. Eur Rev Med Pharmacol Sci, 2020, 24(20):10829-10838. DOI: 10.26355/eurrev_202010_23445.
[5] Skoglund C, Carlsen AL, Weiner M, et al. Circulating microRNA expression pattern separates patients with anti-neutrophil cytoplasmic antibody associated vasculitis from healthy controls[J]. Clin Exp Rheumatol, 2015, 33(2 Suppl 89):S-64-71.
[6] Hu X, Bai S, Li L, et al. MiR-200c-3p increased HDMEC proliferation through the notch signaling pathway[J]. Exp Biol Med (Maywood), 2021, 246(8):897-905. DOI: 10.1177/1535370220981859.
[7] Liu Y, Jiang Y, Li W, et al. MicroRNA-200c-3p inhibits proliferation and migration of renal artery endothelial cells by directly targeting ZEB2[J]. Exp Cell Res, 2020, 387(2):111778. DOI: 10.1016/j.yexcr.2019.111778.
[8] Chen D, Zhang C, Chen J, et al. MiRNA-200c-3p promotes endothelial to mesenchymal transition and neointimal hyperplasia in artery bypass grafts[J]. J Pathol, 2021, 253(2):209-224. DOI: 10.1002/path.5574.
[9] Yun KW, Lee JY, Yun SW, et al. Elevated serum level of microRNA (miRNA)-200c and miRNA-371-5p in children with Kawasaki disease[J]. Pediatr Cardiol, 2014, 35(5):745-752. DOI: 10.1007/s00246-013-0846-6.
[10] Zhang Y, Wang Y. The correlation of plasma microRNA-200 family expressions with risk and disease severity of lupus nephritis[J]. Eur Rev Med Pharmacol Sci, 2018, 22(10):3118-3125. DOI: 10.26355/eurrev_201805_15070.
[11] Senturk T, Antal A, Gunel T. Potential function of microRNAs in thoracic aortic aneurysm and thoracic aortic dissection pathogenesis[J]. Mol Med Rep, 2019, 20(6):5353-5362. DOI: 10.3892/mmr.2019.10761.
[12] Zhang W, Chen P, Zong H, et al. MiR-143-3p targets ATG2B to inhibit autophagy and promote endothelial progenitor cells tube formation in deep vein thrombosis[J]. Tissue Cell, 2020, 67:101453. DOI: 10.1016/j.tice.2020.101453.
[13] Troidl K, Hammerschick T, Albarran-Juarez J, et al. Shear stress-induced miR-143-3p in collateral arteries contributes to outward vessel growth by targeting collagen V-alpha2[J]. Arterioscler Thromb Vasc Biol, 2020, 40(5):e126-e137. DOI: 10.1161/ATVBAHA.120.313316.
[14] Xiang D, Li Y, Cao Y, et al. Different effects of endothelial extracellular vesicles and LPS-induced endothelial extracellular vesicles on vascular smooth muscle cells: role of curcumin and its derivatives[J]. Front Cardiovasc Med, 2021, 8:649352. DOI: 10.3389/fcvm.2021.649352.
[15] Liang H, Chen Y, Li H, et al. MiR-22-3p suppresses endothelial progenitor cell proliferation and migration via inhibiting onecut 1 (OC1)/vascular endothelial growth factor A (VEGFA) signaling pathway and its clinical significance in venous thrombosis[J]. Med Sci Monit, 2020, 26:e925482. DOI: 10.12659/MSM.925482.
[16] Huang SC, Wang M, Wu WB, et al. Mir-22-3p inhibits arterial smooth muscle cell proliferation and migration and neointimal hyperplasia by targeting HMGB1 in arteriosclerosis obliterans[J]. Cell Physiol Biochem, 2017, 42(6):2492-2506. DOI: 10.1159/000480212.
[17] Zhang W, Zhang T, Yan Y, et al. Exosomal miR-22-3p derived from chronic rhinosinusitis with nasal polyps regulates vascular permeability by targeting VE-Cadherin[J]. Biomed Res Int, 2020, 2020:1237678. DOI: 10.1155/2020/1237678.
[18] Takeda E, Suzuki Y, Sato Y. Age-associated downregulation of vasohibin-1 in vascular endothelial cells[J]. Aging Cell, 2016, 15(5):885-892. DOI: 10.1111/acel.12497.
[19] Su YJ, Tsai NW, Kung CT, et al. Investigation of microRNA in mitochondrial apoptotic pathway in systemic lupus erythematosus[J]. Biomed Res Int, 2018, 2018:9026357. DOI: 10.1155/2018/9026357.
[20] Wang X, Chi J, Dong B, et al. MiR-223-3p and miR-22-3p inhibit monosodium urate-induced gouty inflammation by targeting NLRP3[J]. Int J Rheum Dis, 2021, 24(4):599-607. DOI: 10.1111/1756-185X.14089.
[21] Hu H, Wang B, Jiang C, et al. Endothelial progenitor cell-derived exosomes facilitate vascular endothelial cell repair through shuttling miR-21-5p to modulate Thrombospondin-1 expression[J]. Clin Sci (Lond), 2019, 133(14):1629-1644. DOI: 10.1042/CS20190188.
[22] Li Y, Yan L, Zhang W, et al. MicroRNA-21 inhibits platelet-derived growth factor-induced human aortic vascular smooth muscle cell proliferation and migration through targeting activator protein-1[J]. Am J Transl Res, 2014, 6(5):507-516.
[23] Young NA, Valiente GR, Hampton JM, et al. Estrogen-regulated STAT1 activation promotes TLR8 expression to facilitate signaling via microRNA-21 in systemic lupus erythematosus[J]. Clin Immunol, 2017, 176:12-22. DOI: 10.1016/j.clim.2016.12.005.
[24] Dong L, Wang X, Tan J, et al. Decreased expression of microRNA-21 correlates with the imbalance of Th17 and Treg cells in patients with rheumatoid arthritis[J]. J Cell Mol Med, 2014, 18(11):2213-2224. DOI: 10.1111/jcmm.12353.
[25] Lu X, Yu Y, Tan S. The role of the miR-21-5p-mediated inflammatory pathway in ulcerative colitis[J]. Exp Ther Med, 2020, 19(2):981-989. DOI: 10.3892/etm.2019.8277.
[26] Abdallah RT, Keum JS, El-Shewy HM, et al. Plasma kallikrein promotes epidermal growth factor receptor transactivation and signaling in vascular smooth muscle through direct activation of protease-activated receptors[J]. J Biol Chem, 2010, 285(45):35206-35215. DOI: 10.1074/jbc.M110.171769.
[27] Liu J, Feener EP. Plasma kallikrein-kinin system and diabetic retinopathy[J]. Biol Chem, 2013, 394(3):319-328. DOI: 10.1515/hsz-2012-0316.
[28] Xu J, Fang J, Cheng Z, et al. Overexpression of the Kininogen-1 inhibits proliferation and induces apoptosis of glioma cells[J]. J Exp Clin Cancer Res, 2018, 37(1):180. DOI: 10.1186/s13046-018-0833-0.
[29] Vosgerau U, Lauer D, Unger T, et al. Cleaved high molecular weight kininogen, a novel factor in the regulation of matrix metalloproteinases in vascular smooth muscle cells[J]. Biochem Pharmacol, 2010, 79(2):172-179. DOI: 10.1016/j.bcp.2009.08.004.
[30] He X, Yin W, Ding Y, et al. Higher serum angiotensinogen is an indicator of IgA vasculitis with nephritis revealed by comparative proteomes analysis[J]. PLoS One, 2015, 10(6):e0130536. DOI: 10.1371/journal.pone.0130536.
[31] Yu J, Peng R, Chen H, et al. Kininogen 1 and insulin-like growth factor binding protein 6: candidate serum biomarkers of proliferative vitreoretinopathy[J]. Clin Exp Optom, 2014, 97(1):72-79. DOI: 10.1111/cxo.12088.
[32] Liu W, Liu C, Zhang L, et al. Molecular basis of coronary artery dilation and aneurysms in patients with Kawasaki disease based on differential protein expression[J]. Mol Med Rep, 2018, 17(2):2402-2414. DOI: 10.3892/mmr.2017.8111.
[33] Wang LJ, Xue Y, Li H, et al. Wilms′ tumour 1-associating protein inhibits endothelial cell angiogenesis by m6A-dependent epigenetic silencing of desmoplakin in brain arteriovenous malformation[J]. J Cell Mol Med, 2020, 24(9):4981-4991. DOI: 10.1111/jcmm.15101.
[34] Kawasaki A, Namba N, Sada KE, et al. Association of TERT and DSP variants with microscopic polyangiitis and myeloperoxidase-ANCA positive vasculitis in a Japanese population: a genetic association study[J]. Arthritis Res Ther, 2020, 22(1):246. DOI: 10.1186/s13075-020-02347-0.